Особенно высокая активность амилаз и a-глюканфосфорилаз наблюдается при прорастании семян, клубней и луковиц, когда в них происходит интенсивный распад полисахаридов крахмала и увеличивается концентрация декстринов, мальтозы и моносахаридов, используемых для формирования тканей проростков.
В процессе распада крахмал не только может превращаться в мальтозу и глюкозу, но также и в сахарозу. Наиболее активно такие превращения происходят в листьях растений с фотосинтетическим крахмалом. На первом этапе указанных превращений под действием соответствующего трансгликозилирующего фермента остатки глюкозы от крахмала переносятся на УДФ, в результате образуется УДФ-глюкоза:
Затем под действием фермента сахарозо-УДФ-глюкозилтрансферазы остаток глюкозы от УДФ-глюкозы переносится на фруктозу с образованием сахарозы:
УДФ-глюкоза + фруктоза D сахароза + УДФ
Совершенно очевидно, что в результате обращения указанных выше двух реакций возможно также превращение сахарозы в крахмал.
Синтез и распад полифруктозидов. В растениях найдены ферменты, катализирующие превращения сахарозы в полифруктозиды и свободную глюкозу. У некоторых растительных видов (мятликовые, лилейные, астровые, колокольчиковые) образовавшиеся при фотосинтезе углеводы превращаются во фруктозиды, которые затем поступают в созревающие семена или другие запасающие органы. Схематически превращение сахарозы в полифруктозиды можно представить следующим образом:
n сахароза ¾® n глюкоза + [фруктоза]ո
 |
Ферменты, катализирующие указанные превращения содержатся также в клетках бактерий. Донором фруктозных остатков при синтезе полифруктозидов служит УДФ-фруктоза:
При биосинтезе инулина остатки фруктозы присоединяются от УДФ-фруктозы к молекуле сахарозы, включающей остатки глюкозы и фруктозы:
глюкозо-О-фруктоза + УДФ-фруктоза ¾®глюкозо-О-фруктоза-О-фруктоза + УДФ
В ходе реакции между остатками фруктозы образуется b(1®2)-связь. В дальнейшем к полученному продукту вновь осуществляется перенос остатка фруктозы от УДФ-фруктозы с образованием новой b(1®2)-связи. Удлинение полифруктозной цепи по указанному механизму будет продолжаться до тех пор, пока полностью не закончится синтез молекулы инулина. Распад молекулы инулина происходит гидролитическим путём под действием фермента инулазы (3.2.1.7) с образованием свободной фруктозы и сахарозы. Другие полифруктозиды гидролизуются соответствую-щими гликозидазами до свободной фруктозы.
Синтез и распад целлюлозы. Молекулы целлюлозы построены из остатков b-D-глюкозы, соединённых b(1®4)-связями. В опытах in vitro с использованием бесклеточных ферментных экстрактов, выделенных из растений, показано, что синтез клетчатки может осуществляться из УДФ-глюкозы или ГДФ-глюкозы (гуанозиндифосфатглюкозы), а также фрагментов целлюлозы (целлодекстринов), выполняющих роль акцепторов остатков b-глюкозы.
Налог на доходы физических лиц. Платим налог и получаем вычеты
Содержащийся в составе этих экстрактов фермент целлюлозосинтетаза (2.7.7.34) катализирует синтез молекул целлюлозы по схеме:
Молекулы фермента образуют полиферментный комплекс, который катализирует одновременный синтез многих молекул целлюлозы, формирующих микрофибриллу. В микрофибриллах полимерные цепи из остатков β-глюкозы соединены водородными связями.
Молекулы целлюлозы расщепляются в основном путём гидролиза под действием фермента целлюлазы (3.2.1.4) с образованием целлобиозы. Особенно высокой целлюлазной активностью отличаются клетки многих почвенных микроорганизмов, а также клетки бактерий пищеварительной системы жвачных животных. Имеются также сведения о том, что и в листьях растений при их старении происходит процесс распада целлюлозы и превращения её в растворимые формы, которые затем подвергаются оттоку в созревающее зерно или клубни.
Продукт гидролиза целлюлозы – целлобиоза подвергается гидролитическому расщеплению с участием фермента b-глюкозидазы (3.2.1.21) до свободной b-глюкозы.
Гемицеллюлозы. В биосинтезе гемицеллюлоз также принимают участие ферменты, катализирующие перенос остатков моносахаридов от их нуклиозиддифосфатпроизводных на соответствующий олигосахарид, служащий первичным акцептором при образовании полисахарида. В качестве донора остатков ксилозы для синтеза ксиланов используется УДФ-ксилоза. Донором остатков арабинозы при образовании молекул арабанов служит УДФ-арабиноза. Для синтеза маннанов активированной формой моносахаридных остатков для включения в состав полисахаридной цепи является ГДФ-манноза (гуанозиндифосфатманноза).
Распад гемицеллюлоз происходит в основном гидролитическим путём под действием ферментов гемицеллюлаз. Такие ферменты найдены в клетках бактерий и плесневых грибов, а также в прорастающих семенах растений. Активирование гемицеллюлаз наблюдается в листьях растений в процессе их старения, при этом образующиеся в результате гидролиза гемицеллюлоз растворимые формы углеводов поступают в созревающие семена или клубни (у картофеля). Отмечается распад гемицеллюлоз и при созревании плодов у некоторых плодовых культур.
Пектиновые вещества. Пектины синтезируются из УДФ-галактуроновой кислоты, которая образуется при окислении УДФ-глюкозы или УДФ-галактозы, а также в результате активирования свободной галактуроновой кислоты путём фосфорилирования и взаимодействия с УТФ:
галактуроновая кислота + АТФ ¾® 1-фосфогалактуронова кислота + АДФ
1-фосфогалактуроновая кислота + УТФ ¾® УДФ-галактуроновая кислота + Н4Р2О7

В ходе образования пектинов остатки a- D -галактуроновой кислоты под действием специфических трансфераз переносятся от УДФ-галактуроновой кислоты на соответствующий акцептор. Одновременно происходят реакции метилирования карбоксильных групп под действием фермента метилтрансферазы, катализирующего перенос метильных радикалов от S-аденозилметионина на полигалактуроновую кислоту:
При взаимодействии пектина с галактанами и арабанами осуществляется синтез сложноэфирных связей протопектина с участием карбоксильных групп, содержащихся в молекулах пектина. Эти реакции также катализируют специфические трансферазы, осуществляющие синтез протопектина в клеточных стенках растений или в формирующихся плодах.
Распад протопекина происходит гидролитическим путём под действием ферментов протопектиназ, катализирующих гидролиз сложноэфирных связей, соединяющих полимерные цепи пектина с галактанами и арабанами. Процессы гидролиза протопекинов очень активно протекают в растениях при созревании плодов и старении листьев.
Под действием фермента пектазы (3.1.1.11), катализирующего гидролиз сложноэфирных связей метоксильных групп, пектины превращаются в полигалактуроновую (пектиновую) кислоту и метиловый спирт. Полигалактуроновая кислота в отличие от пектина не обладает способностью образовывать желе, но очень легко взаимодействует с катионами металлов, образуя нерастворимые соли. На этом основано химическое определение пектиновых веществ и происходит их защитное действие в пищеварительной системе человека и животных по связыванию и выведению из организма катионов тяжелых металлов.
Пектиновая кислота подвергается гидролизу с участием фермента полигалактуроназы (3.2.1.15), или пектиназы, действующей на О-глико-зидные связи, соединяющие остатки a- D -галактуроновой кислоты. Комплекс ферментов, гидролизующих пектиновые вещества, особенно активен в препаратах, получаемых из плесневых грибов. В связи с этим указанные препараты применяются для осветления плодовых соков и вин, содержащих растворимый пектин, который является причиной их возможной непрозрачности.
Вопросы для повторения:
1. В какой последовательности происходит синтез гексозы из СО2 и Н2О в ходе реакций цикла Кальвина? 2. При каких условиях усиливаются реакции фотодыхания и как они влияют на эффективность процесса фотосинтеза? 3. Как происходит ассимиляция СО2 у С4-растений? 4. Какие биохимические реакции происходят в пентозофосфатном цикле и какова его биологическая роль?
5. Как происходит синтез из моносахаридов спиртов? 6. Какие известны взаимные превращения гексоз (глюкозы, фруктозы, маннозы и галактозы)? 7. Каковы механизмы образования в организмах рибозы, ксилозы, арабинозы, эритрозы, глицеринового альдегида и диоксиацетона? 8. Как осуществляются синтез и превращения сахарозы и других олигосахаридов?
9. Какие биохимические реакции лежат в основе синтеза и распада крахмала, полифруктозидов, целлюлозы, гемицеллюлоз, пектиновых веществ? 10. С участием каких ферментов происходят синтез, превращения и распад раличных моносахаридов, сахарозы, крахмала и других полисахаридов?
Резюме по модульной единице. Первичный синтез углеводов в фотосинте-зирующих клетках осуществляется в ходе реакций цикла Кальвина. У С₃-растений эти реакции происходят в тех же клетках, в которых протекают фотохимические реакции усвоения солнечной энергии, а у С₄-растений – в клетках обкладочной паренхимы.
В реакциях цикла Кальвина образуются не свободные моносахариды, а их фосфорнокислые эфиры – 3-фосфоглицериновый альдегид, фосфодиоксиацетон, эритрозо-4-фосфат, рибозо-5-фосфат, фруктозо-6-фосфат. В нефотосинтезирующих клетках указанные фосфаты моносахаридов синтезируются в ходе реакций пентозофосфатного цикла.
Фосфорнокислые эфиры триоз образуются также в анаэробной стадии дыхания. Фосфаты моносахаридов легко подвергаются взаимопревращениям под действием соответствующих изомераз. Полный распад моносахаридов осуществляется в реакциях дыхания. Ксилоза и арабиноза образуются в результате окислительного декарбоксилирования глюкозы и галактозы.
В синтезе олиго- и полисахаридов важную роль играют нуклеозиддифосфат-производные моносахаридов – УДФ-глюкоза, УДФ-фруктоза, ГДФ-глюкоза, АДФ-глюкоза, УДФ-галактуроновая кислота и др. Они служат донорами моносахаридных остатков при синтезе олиго- и полисахаридов. Активирование молекул моносахаридов происходит путём фосфорилирования, которое катализируют ферменты – киназы (фосфотрансферазы). Синтез сложных углеводов катализируют гликозилтрансферазы, а распад – соответствующие гликозидазы – β-фруктофуранозидаза, α-глюкозидаза, амилазы, инулаза, протопектиназы и пектиназы, целлюлазы и др. В распаде крахмала участвуют также фосфорилазы, под действием которых из крахмала образуется глюкозо-1-фосфат.
Тестовые задания к лекции. Тесты № 115-155.
Аннотация. Излагаются механизмы синтеза, превращений и распада основных групп липидов в организмах. Даются особенности действия ферментов и ферментных комплексов, катализирующих эти биохимические реакции. Указывается локализация реакций синтеза и превращений конкретных липидов. Рассматриваются биохимические механизмы превращения липидов в углеводы и включения продуктов распада липидов в дыхательные реакции.
Ключевые слова. Синтетаза жирных кислот, ацилпереносящие белки, ацетил-кофермент А, пропионилкофермент А, малонилкофермент А, метилмалонилкофермент А, оксигеназы (десатуразы), глицеролфосфатацилтрансферазы, ЦДФ-этаноламин, ЦДФ-холин, липазы, α-окисление жирных кислот, β-окисление жирных кислот, карнитин, глиоксилатный цикл, глиоксисомы, глюконеогенез, фосфолипазы, циклоартенол.
Рассматриваемые вопросы:
1. Синтез глицеролфосфата и жирных кислот.
2. Синтез ацилглицеринов и фосфолипидов.
3. Распад жиров и фосфолипидов.
4. Превращение жирных кислот в углеводы.
5. Синтез и превращения других липидов.
Понравилась статья? Добавь ее в закладку (CTRL+D) и не забудь поделиться с друзьями:
Источник: studopedia.ru
Синтез триацилглицеролов

Образовавшиеся глицерол-3-фосфат и ацил-КоА жирных кислот – исходные соединения для биосинтеза жира (см. рис. 11, Этап V).
На первом этапе синтеза триацилглицеролов образуется 3-фосфатидная кислота (диацилглицерол-3-фосфат), при этом ацильные остатки КоА-производных жирных кислот переносятся специфическими ацил-трансферазами на спиртовые группы глицерол-3-фосфата:
На втором этапе фосфатидная кислота гидролизуется фосфатидатфосфатазой с образованием 1,2-диацилглицерола:
Затем 1,2-диацилглицерол превращается в триацилглицерол путем переноса остатка КоА-производного третьей жирной кислоты. Этот перенос катализируется ферментом диацилглицеролацилтрансферазой:
3 Метаболизм фосфолипидов
Распад (катаболизм) фосфолипидов
Фосфолипиды также как и жиры, активно распадаются в тканях животных и растений.
Гидролиз фосфолипидов осуществляют несколько фосфолипаз, различающихся по расщеплению определенных связей и получивших название фосфолипаз А, С и Д:
Здесь R1 и R2 – углеводородные радикалы; В – остатки азотсодержащих одноатомных спиртов (холина, этаноламина, серина).
Фосфолипаза А1 отщепляет жирные кислоты по α-положению; фосфолипаза А2 осуществляет ту же самую реакцию по β-положению; фосфолипаза С отщепляет фосфорилированные азотсодержащие спирты от фосфолипидов; фосфолипаза D гидролизует фосфолипиды с образованием азотсодержащего спирта и фосфатидной кислоты.
В растениях обнаружены все виды расщепления. Фосфолипазы А обнаружены также у животных и микроорганизмов. Фосфолипазы локализованы преимущественно в лизосомах. Конечными продуктами гидролиза фосфолипидов является глицерин, жирные кислоты, азотистые спирты и фосфорная кислота.
Синтез фосфолипидов
В отличие от триацилглицеролов и жирных кислот фосфолипиды не являются существенным энергетическим материалом. Фосфолипиды, как было сказано ранее, играют важную роль в структуре и функции клеточных мембран.
Биосинтез фосфолипидов интенсивно происходит в печени, стенке кишечника, молочной железе и других тканях. Наиболее важные фосфолипиды синтезируются главным образом в эндоплазматической сетке клетки.
Центральную роль в биосинтезе фосфолипидов играют L-диацилглицеролы (в синтезе фосфатидилхолинов и фосфатидилэтаноламинов), фосфатидная кислота (в синтезе фосфатидилинозитов) и сфингозин (в синтезе сфингомиелинов).
В этих биосинтезах принимает участие цитидинтрифосфат (ЦТФ), выполняющий роль активатора.
Основными компонентами мембран являются фосфатидилэтаноламин и фосфатидилхолин.
В процессе синтеза фосфатидилэтаноламина вначале происходит активация этаноамина под действием этаноламинкиназы:
Этаноламинфосфат реагирует затем с цитидинтрифосфатом (ЦТФ), в результате чего образуется цитидиндифосфатэтаноламин (ЦДФ-этаноламин) и пирофосфат (РРі). Эту реакцию катализирует фосфоэтаноламинцитидинтрансфераза:
Активированный ЦДФ–этаноламин при участии фермента этаноламинфосфаттрансферазы вступает в реакцию с L–диацилглицеролом с образованием фосфатидиламина:
L-диацилглицерол образуется при гидролизе фосфатидной кислоты. Аналогичные реакции ведут к образованию ЦДФ-холина.; последний вступает в реакцию с L-диацилглицеролом, в результате чего образуется фосфатидилхолин.
В синтезе фосфатидилэтаноламина и фосфатидилхолина важную роль играет ЦТФ. Подобно тому синтезу олиго- и полисахаридов, мы и здесь видим, каким образом нуклеотиды могут выполнять функцию переносчиков опреденных химических групп в обмене веществ клеток.
Помимо синтеза фосфолипидов таким способом, в клетке они легко подвергаются взаимопревращениям. Фосфатидилэтаноламин является предшественником фосфатидилхолина. Так, в печени в результате последовательного переноса трех метильных групп от трех молекул S-аденозилметионина образуется фосфатидилхолин:
В переносе метильных групп важную роль играют тетрогидрофолевая кислота (витамин Вс) и метилкобаломин ( витамин В12).
Синтез фосфатидилсерина происходит другим путем с предварительным вовлечением фосфатидной кислоты:
Затем происходит перенос серина на фосфатидильный остаток с образованием фосфатидилсерина:
У млекопитающих фосфатидилсерин образуется в реакции обмена этаноламина на серин следующим путем:
Таким же путем образуется фосфатидилинозитол.
В биосинтезе сфингомиелина принимает участие церамид (N-ацетилсфингозин), который образуется при взаимодействии спирта сфингозина и ацетил-КоА. Сфингомиелин синтезируется в результате взаимодействия церамида с ЦДФ-холином:
Все выше синтезированные фосфолипиды поступают с помощью липидпереносящих белков из цитоплазмы к мембранам и участвуют в их построении.
Источник: megaobuchalka.ru
2.1 Свойства моносахаридов
Моносахариды, являясь альдо- или кетоспиртами, проявляют все свойства альдегидов, кетонов и спиртов.
Образование гликозидов
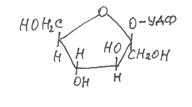
В результате взаимодействия гликозидного гидроксила моносахарида со спиртами, образуются соединения типа простых эфиров, которые называются гликозидами:
Вновь образовавшаяся связь называется гликозидной.
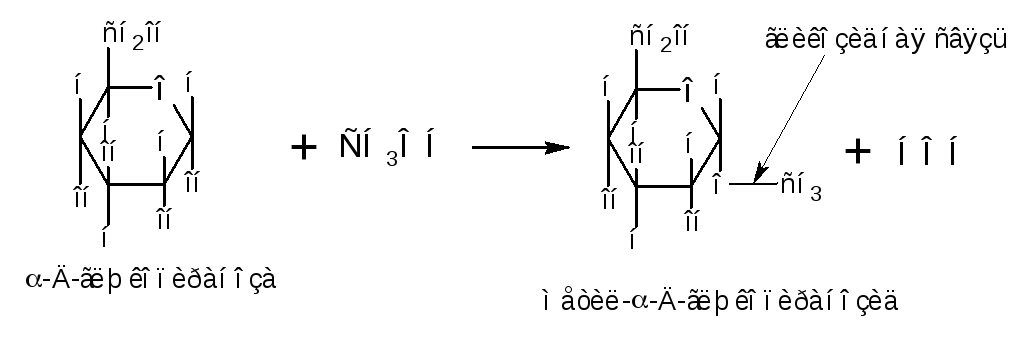
Гликозидная связь имеет очень важное биологическое значение. С помощью этой связи осуществляется ковалентное связывание моносахаридов в составе олиго- и полисахаридов:
В природе встречаются все виды гликозидных связей:13;12;16; но каждый конкретный олиго- или полисахарид содержит определенный вид гликозидных связей между мономерными единицами. Кроме О-гликозидов в природе встречаютсяS-гликозиды (тиогликозиды) иN-гликозиды (нитрилгликозиды). Гликозиды широко распространены в растениях, они часто обладают специфическим запахом и вкусом.
Например, гликозид синигрин, содержится в семенах горчицы, при гидролизе образует эфирное горчичное масло, придающее специфический вкус и аромат пищевой горчице. Гликозид глюкованилин синтезируется в ванильном дереве, амигдалин содержится в семенах миндаля, вишен, персиков.
Для гидролитического расщепления гликозидов широко применяется ферментативный гидролиз, преимущество которго заключается в его специфичности. Например, фермент -глюкозидаза из дрожжей расщепляет только-гликозидную связь;-глюкозидаза из миндаля – только-гликозидную связь. На этом основании ферментативный гидролиз часто применяется в целях установления конфигурации аномерного атома углерода. Гидролиз гликозидов лежит в основе гидролитического расщепления полисахаридов, осуществляемого в организме, а также используется во многих промышленных процессах.
2 .Образование сложных эфиров
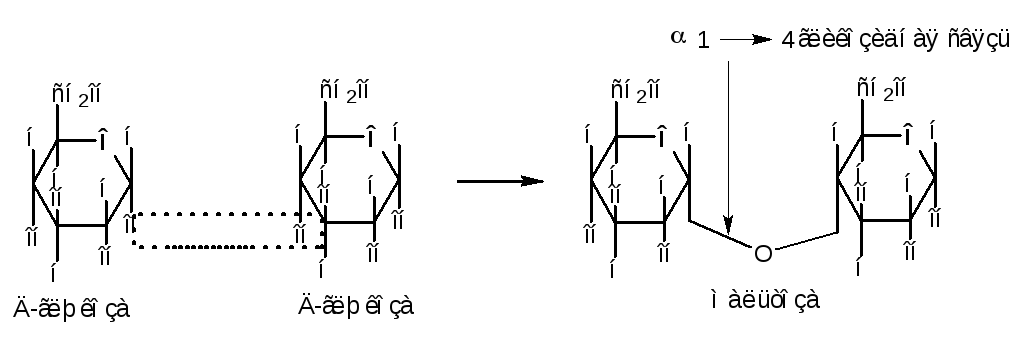
Моносахариды, реагируя с кислотами, могут давать сложные эфиры. Из производных сахаров наиболее важное биологическое значение имеют эфиры фосфорной кислоты.
В клетке эти производные легко образуются при ферментативных реакциях:
Наиболее важную роль в превращениях крахмала, а также в процессах дыхания и спиртового брожения играют глюкозо-6-фосфат, глюкозо-1-фосфат, фруктозо-6-фосфат, фруктозо-1,6-дифосфат. Донором фосфорной кислоты является АТФ, в молекуле которой расщепляется высокоэнергетическая связь. Биологическое значение сахарофосфатов состоит в том, что они представляют собой метаболически активные формы сахаров, играющие важную роль в биоэнергетике живой клетки.
Фосфорилирование моносахаридов переводит их в мобильное реакционно-способное состояние, а при их биологическом окислении химическая энергия моносахарида аккумулируется в фосфатном радикале.
Фосфорилирование выгодно клетке еще и потому, что клеточная мембрана малопроницаема для фосфорных эфиров моносахаридов, что очень важно для активного транспорта моносахаридов из внешней среды внутрь клетки.
3. Образование нуклеозиддифосфатсахаров
В биосинтезе олиго- и полисахаридов при взаимопревращениях моносахаридов важную роль играют нуклеозиддифосфатсахара (уридиндифосфаты).
Источник: studfile.net